
Rozmnožovanie nahosemenných rastlín
Nahosemenné rastliny, známe aj ako gymnospermy, predstavujú významnú skupinu semenných rastlín s charakteristickým spôsobom rozmnožovania, ktorý ich odlišuje od krytosemenných rastlín. Ich evolučný vývoj, ktorý siaha do obdobia karbónu, ich zaraďuje k prechodným formám medzi výtrusnými a krytosemennými rastlinami.
Charakteristika nahosemenných rastlín
Hlavným znakom nahosemenných rastlín je tvorba semien, ktoré však nie sú ukryté v semenníkoch, ale ležia voľne na semenných šupinách. Preto sa nazývajú „nahé“. Tieto rastliny nemajú kvety v pravom zmysle slova, s obalmi a bliznami. Namiesto toho tvoria jednopohlavné samčie a samičie šištice (strobilus).
Z morfologického hľadiska je plod rozmnožovací orgán výlučne krytosemenných rastlín. Vzniká zo steny semenníka po oplodnení vajíčka. Nahosemenné rastliny (ako borovice, cyprusy, ginko, tis) však nemajú semenníky. Keďže nemajú semenník, po oplodnení vajíčka z neho vznikne len nahé semeno bez oplodia - a teda z definície nemôže vzniknúť plod.
Mikrosporofyly (tyčinky) nahosemenných rastlín sú zoskupené do šištíc (strobily) a môžeme ich považovať za jednopohlavný kvet. Tyčinky sa skladajú z nitky a mikrosporangia (peľnice). Z peľnice sa uvoľňujú mikrospóry (t. j. peľové zrná). Makrosporofyly sa skladajú zo sterilných šupín, ktoré môžu byť rozlíšené na podporné a semenné. Megaspórou je vajíčko (ovulum), ktoré je v čase zrelosti viacbunkové.
Špecifická je aj stavba ich dreva. Vodivé pletivá (xylém) sú jednoduchšie - namiesto pravých ciev obsahujú len cievice (tracheidy), čo z nich robí takzvané mäkké drevo. Mnohé druhy si pletivá chránia tvorbou živice.
Rozmnožovanie
Rozmnožovanie nahosemenných rastlín je proces, ktorý zahŕňa tvorbu samčích a samičích gamét a ich následné spojenie. Tento proces je prispôsobený prostrediu a často prebieha s pomocou vetra.
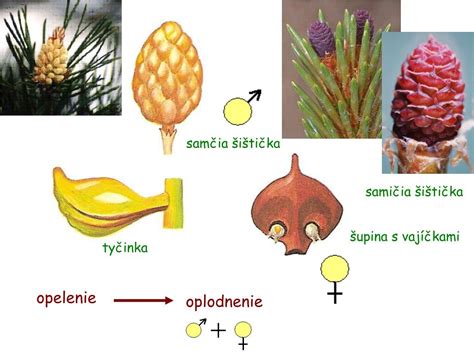
Samčia šištica (mikrostrobilus)
Samčia šištica je tvorená krátkym vretenom a blanitými, špirálovite usporiadanými šupinami priliehajúcimi k vretenu. Vyrastajú v spodných etážach stromu. Blanitá šupina - mikrosporofyl - predstavuje tyčinku s dvoma alebo niekoľkými mikrosporangiami. Vývin samčieho gametofytu prebieha vo vnútri peľového zrna. Samčie šištice vyrastajú spravidla na jar, na strome ostávajú niekoľko týždňov a potom opadávajú.
Vo vnútri juvenilného mikrosporangia sa nachádza peľotvorné pletivo - archespór. Z neho sa diferencuje výstelkové pletivo - tapetum a sporogénne pletivo. Redukčným delením buniek sporogénneho pletiva vzniknú tetrády haploidných mikrospór. Bunková stena vyvíjajúcich sa peľových zŕn je v tomto období tvorená exinou a vnútornou intinou. U borovice a ďalších druhov ihličnatých rastlín exina odstáva a vytvárajú sa vzdušné vaky, ktoré umožňujú prenos peľových zŕn na veľké vzdialenosti.
Mikrospóry, podobne ako megaspóry, sa začínajú deliť ešte v sporangiu. Vo vnútri peľového zrna sa v jednom mieste v blízkosti bunkovej blany oddelí niekoľko drobných buniek, ktoré väčšinou po určitom čase zanikajú. Tieto bunky predstavujú samčie prothálium. Inak vnútro peľového zrna vypĺňa veľká bunka, ktorá sa mitoticky delí. Po celularizácii vzniká väčšia vegetatívna bunka a menšia generatívna bunka. Generatívna bunka sa opäť delí a vznikne bunka nástenná, ktorá susedí s protháliovými bunkami a spermatogénna bunka, z ktorej pri prerastaní peľového vrecúška vzniknú 2 spermatické bunky. U primitívnejších skupín nahosemenných rastlín (napr. cykasovitých) sa samčie gaméty vyvíjajú ako obrvené spermatozoidy.
Samičia šištica (megastrobilus)
Samičia šištica je tvorená zdrevnateným vretenom, na ktorom sú špirálovite usporiadané podporné šupiny a v ich úžľabí vyrastajú semenné šupiny. U borovice sú podporné šupiny zakrpatené a semennými šupinami prerastené natoľko, že sú morfologicky takmer nerozoznateľné. Na vrchnej strane semenných šupín sú pri báze lokalizované dve vajíčka.
Vajíčka nahosemenných rastlín sú krasinucelátne. Nucelus je pokrytý výraznou pokožkou, ktorá je obklopená veľkým počtom parenchymatických buniek. Na vrchole je nucelus sformovaný do peľovej komory, ktorá vzniká buď tak, že skupina buniek v apikálnej časti lyzuje a vzniká plytká priehlbenina, alebo sa nápadne naddvihne a vzniká poduškovitý útvar. Oba tieto útvary slúžia na vytvorenie vhodných podmienok pre zachytenie a vyklíčenie peľového zrna. Integument postupne prerastá nucelus a vytvorí sa mikropyla, ktorá podobne ako blizna krytosemenných rastlín, produkuje polinačnú kvapku.
Sporogénne pletivo sa zakladá z jednej alebo viacerých subepidermálnych buniek nucela. Vzniká primárny archespór. Bunky primárneho archespóru sa delia najprv antiklimálne a neskôr periklinálne. Vzniká vonkajšia - parietálna a vnútorná sporogénna vrstva, teda sekundárny archespór. V sekundárnom archespóre prebieha ešte opakované mitotické delenie buniek. Z týchto buniek sa niektoré transformujú na megasporocyty.
Megasporocyt sa redukčne delí a vzniká tetráda haploidných megaspór. Pri niektorých taxónoch boli pozorované iba tri megaspóry. K tomu dochádza vtedy, ak jedna z dcérskych buniek, ktoré vznikli pri heterotypickom delení sa ďalej nedelí. Býva to najčastejšie bunka, ktorá sa nachádza najbližšie pri mikropyle. Postavenie megaspór je zvyčajne lineárne, ale môže byť aj tetraedrické alebo postavenie v tvare T. U Pinus silvestris v procese heterotypického delenia vznikajú dve, nerovnako veľké bunky. Väčšia je uložená v chalazálnej oblasti, menšia v mikropilárnej. Ďalej sa vyvíja len bazálna megaspóra, ktorej niekoľkonásobným mitotickými delením vzniká voľnojadrové prothálium. Výskyt veľkého počtu voľných jadier je charakteristickým znakom všetkých nahosemenných rastlín. V ďalšom procese sa vytvoria bunkové steny. Z týchto haploidných buniek vzniká endosperm a archegóniá. Iniciálna bunka archegónia sa delí a vzniká vonkajšia tzv. primárna hrdlová bunka a vnútorná tzv. primárna brušná bunka. Delením primárnej hrdlovej bunky vzniká hrdlo archegónia a delením primárnej brušnej bunky vzniká brušná kanálová bunka a oosféra. Samičí gametofyt Pinus silvestris teda pozostáva z haploidného endospermu a z dvoch archegónií umiestnených v oblasti mikropily. V každom archegóniu je jedna oosféra. Endospermové jadrá sú haploidné, pri celularizácii sa však môžu dostať do jednej bunky aj viaceré jadrá, ktoré môžu splývať a vytvárať polyploidné endospermálne pletivo. Endosperm má charakter zásobného parenchýmu a obsahuje predovšetkým škrob, aleuronové zrná a lipidy.
Opelenie a oplodnenie
Počas prvej sezóny sa šupiny samičích šištíc so zrelými vajíčkami oddialia od vretena. Peľové zrná dopadajú medzi šupiny a na povrchu vajíčka sa zachytávajú do polinačnej kvapky, ktorá je vylučovaná v oblasti mikropily. Pri vysýchaní kvapky sa peľové zrno dostane na povrch nucela. Peľové zrno klíči, vzniká peľové vrecúško, ktoré začne prerastať cez nucelus. Šupiny samičích šištíc sa v tomto období primknú k vretenu. Spermatogénna bunka sa v peľovom vrecúšku rozdelí na dve spermatické bunky. Jedna spermatická bunka oplodní oosféru - vzniká zygota. Ostatné bunky peľového vrecúška zanikajú. Oplodnená býva zvyčajne iba jedna vajcová bunka.
Opeľuje ich prevažne vietor. Vytvárajú peľovú trubicu, čím sa pri oplodnení úplne oslobodili od vody. Peľ prechádza priamo k vajíčku a oplodnenie zabezpečuje len jedna spermatická bunka.
Vývin embrya a vznik semena
Zo zygoty sa ďalším delením a diferenciáciou vyvíja embryo a celé vajíčko sa premieňa na semeno. Z vnútornej vrstvy integumentu vzniká osemenie a z vonkajšej vrstvy sa diferencuje krídielko, ktoré napomáha rozširovaniu semien vetrom.
V procese embryogenézy je charakteristický jav voľnojadrového delenia, to znamená, že v počiatočných fázach sa jadro zygoty delí, ale nevznikajú bunkové steny. Počet voľnojadrových delení je u jednotlivých rodov rôzny a až potom nastáva celularizácia. Napríklad u Ginkgo biloba nastáva cytokinéza až po vytvorení 256 jadier, u cykasu Dioon edule 1000 a u ihličnanov po vytvorení 2 až 64 jadier.
Ďalšou zvláštnosťou ihličnatých rastlín je polyembryónia. Napríklad v rode Pinus v prvej fáze voľnojadrového delenia vzniknú štyri jadrá, ktoré putujú k mikropyle a usporiadajú sa do jednej vrstvy. Jadrá sa delia, vzniká druhá vrstva jadier a súčasne sa diferencujú aj bunkové steny. Bunky sa opäť delia a vznikajú štyri vrstvy. Štyri horné - bazálne bunky, pod nimi sú ružicovité (rozetové) bunky a štyri primárne suspenzorové bunky. Na tieto sa pripájajú štyri bunky označované ako iniciály zárodkov. Z nich sa postupne vyvíjajú embryá. Pri niektorých druhoch sa rozetové bunky opätovne delia, vznikajú bočné (sekundárne) suspenzorové bunky zakončené iniciálami embryií.
Zástupcovia a ich rozmnožovanie
Nahosemenné rastliny sú zastúpené rôznymi skupinami, z ktorých každá má svoje špecifiká v rozmnožovaní:
- Cykasorasty (Cycadophyta): Sú striktne dvojdomé a na rozdiel od iných nahosemenných rastlín ich opeľuje hmyz (najmä chrobáky). Vytvárajú semená s nápadným mäsitým obalom (sarkotestou), ktoré vzhľadom pripomínajú kôstkovicu.
- Ginkorasty (Ginkgophyta): Najznámejším zástupcom je Ginkgo biloba. Ide o dvojdomú rastlinu s charakteristickými vejárovitými listami. Netvorí klasické šišky, ale mäsité semená s tvrdou kôstkou (pripomínajúce plody), ktoré po dozretí silno zapáchajú.
- Borovicorasty (Pinophyta): Najpočetnejšia skupina, do ktorej patria ihličnany. Zvyčajne ide o jednodomé rastliny tvoriace drevnaté šišky. Napríklad Pinus silvestris (borovica lesná) má samčie a samičie šištice. Samičie šištice vyrastajú na vrchných konároch.
- Tis obyčajný (Taxus baccata): Je to dvojdomá rastlina, ktorej semená sú chránené červeným sladkým mieškom.
Niektoré ihličnany nemajú drevnaté šištice a nie sú ani jednodomé. Napríklad tis (Taxus baccata) produkuje vajíčka na koncoch krátkych axilárnych výhonkov v mäsitých pohárikovitých arilusoch. Tis je dvojdomá rastlina.
Porovnanie s krytosemennými rastlinami
Nahosemenné rastliny tvoria evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Zatiaľ čo u krytosemenných rastlín je samičí gametofyt zredukovaný len na 7 buniek (zárodočný vak) a samčí gametofyt dokonca len na 2-3 bunky (peľové zrnko), u nahosemenných rastlín je tento proces odlišný. Kým u krytosemenných rastlín dochádza k dvojitému oplodneniu, u nahosemenných rastlín oplodnenie zabezpečuje len jedna spermatická bunka.
Vegetatívne sa rozmnožujú len zriedka, zatiaľ čo ich hlavnou stratégiou rozmnožovania je tvorba semien. Pri pohlavnom rozmnožovaní sa nová rastlina vyvinie zo zygoty, ktorá vznikla splynutím dvoch diferencovaných pohlavných buniek, gamét - samčej gaméty spermatozoidu, pri kvitnúcich rastlinách peľového zrnka, so samičou gamétou vajcovou bunkou.
tags: #rozmnozovanie #nahosemennych #rastlin #ppt