
Rozmnožovanie semenných rastlín: Borovicorasty a Magnóliorasty
Vyššie rastliny (Cormobionta, Embryobionta) sa vyvinuli pred viac ako 500 miliónmi rokov zo zelených rias, konkrétne zo skupiny chary. Sú to prevažne autotrofné rastliny, v chloroplastoch ktorých sa nachádza dvojica chlorofylov a + b. Počas evolúcie urobili kľúčový krok - opustili vodné prostredie a prispôsobili sa životu na suchej zemi. Na rozdiel od nediferencovanej stielky nižších rastlín má telo vyšších rastlín zložitejšiu stavbu a nazýva sa kormus. Prvé suchozemské rastliny sa museli chrániť pred vysušením, čo vyriešili vznikom pokožky. Keďže rastlina už nebola celá ponorená vo vode, musela vybudovať systém rozvozu vody a v nej rozpustených látok po celom tele. Vytvorili sa tak diferencované cievne zväzky (vodivé pletivá). Telo sa skladá z diferencovaných buniek, ktoré vytvárajú pletivá a pravé orgány so špecifickými funkciami: koreň - vyšpecializoval sa na príjem vodných roztokov a upevnenie rastliny k podkladu, stonka - prispôsobila sa na rozvoz živín a primerané rozmiestnenie rastlinných orgánov v priestore, listy - skoncentrovala sa v nich fotosyntéza, pričom rozšírenie listu do plochy ju urobilo oveľa efektívnejšou. Procesy spojené s vytváraním pohlavných buniek a rozmnožovaním sa taktiež situovali do špecifických orgánov.
Semenné rastliny (Spermatophyta) dosiahli evolučný triumf, pretože sa pri oplodnení úplne oslobodili od vody. Ich samčie bunky (spermácie) už zväčša nemajú bičíky. Namiesto plávania vytvára rastlina peľovú trubicu, ktorá bunky bezpečne dopraví priamo k vajíčku. Tieto rastliny už neuvoľňujú spóry do prostredia; ich gametofyt je extrémne zredukovaný a vyvíja sa chránený hlboko vo vnútri materskej rastliny (endospóricky). Hlavným znakom semenných rastlín je to, že vytvárajú kvet (flos), v ktorom sa nachádzajú pohlavné orgány (tyčinky, piestiky) produkujúce peľ a vajíčka. Kvet predstavuje de facto jediný gametofytický útvar. Výsledkom rozmnožovania je mnohobunkové semeno, ktoré chráni embryo pred vyschnutím a obsahuje preň zásobu živín.
Nahosemenné rastliny (Gymnospermae)
Nahosemenné rastliny (iné názvy: nahosemenné, gymnospermy; lat. Gymnospermae) sú veľký taxón semenných rastlín. V mnohých systémoch sa namiesto nich uvádzali len ich jednotlivé časti ako samostatné taxóny. Avšak najnovšie molekulárne poznatky naznačujú, že nahosemenné rastliny spolu s krytosemennými rastlinami tvoria dve monofyletické vetvy, čo viedlo k návratu k pôvodnej koncepcii považujúcej skupinu Gymnospermophyta za monofyletickú. Nahosemenné rastliny predstavujú evolučný prechod medzi výtrusnými cievnatými a krytosemennými rastlinami. Vyvinuli sa koncom starších prvohôr, kedy vplyvom suchej klímy získali malé, kožovité listy s hrubou kutikulou. Dnešné druhy majú listy zväčša ihlicovité alebo šupinovité a sú prevažne vždyzelené.
Najdôležitejším znakom nahosemenných rastlín je tvorba semien. Vajíčka a z nich vzniknuté semená nie sú ukryté v semenníku (netvoria plody), ale ležia voľne exponované na semenných šupinách, preto sa nazývajú „nahé“. Chýbajú im typické kvety s obalmi a bliznou. Kvety sa zoskupujú do jednopohlavných samčích a samičích šišiek (strobilov). Je dôležité zdôrazniť, že termín „plod“ sa v botanickom zmysle vzťahuje výlučne na krytosemenné rastliny, pretože vzniká zo steny semenníka. Nahosemenné rastliny, ktoré nemajú semenník, z neho nemôžu vyvinúť plod. Opeľovanie u nahosemenných rastlín prebieha prevažne pomocou vetra. Vytvárajú peľovú trubicu, čím sa pri oplodnení oslobodili od vody. Peľ prechádza priamo k vajíčku a oplodnenie zabezpečuje len jedna spermatická bunka. Špecifická je aj stavba ich dreva. Vodivé pletivá (xylém) sú jednoduchšie - namiesto pravých ciev obsahujú len cievice (tracheidy), čo im dáva charakter mäkkého dreva. Mnohé druhy si pletivá chránia tvorbou živice.
V starších systémoch sa všetky recentné druhy zaraďovali do oddelenia borovicorastov (Pinophyta), ktoré malo dve hlavné vývojové vetvy: ihličnany a ginká.

Borovicorasty (Pinophyta)
Borovicorasty (iné názvy: borovicorasty, ihličinorasty; lat. Pinophyta, Coniferophyta) sú veľké oddelenie patriace do nahosemenných rastlín. Ihličnany (iné názvy: ihličiny, konifery) v taxonomickom zmysle sú veľká trieda (prípadne neformálna skupina) patriaca do nahosemenných rastlín. Borovicorasty tvoria najpočetnejšiu a evolučne najúspešnejšiu skupinu nahosemenných rastlín (asi 550 druhov). Borovicorasty sú dreviny, väčšinou stromy, málokedy kríky (napr. kosodrevina). Patria k nim najväčšie stromy na svete, sekvoja vždyzelená (Sequoia sempervirens) a sekvojovec mamutí (Sequoiadendron giganteum), ktoré rastú v oblasti západného pobrežia Severnej Ameriky. Borovicorasty majú väčšinou silnú apikálnu dominanciu s monopodiálnym rozkonárovaním, ktoré tvorí viac-menej pravidelné prasleny bočných konárov. Kmene borovicorastov majú kambiálne kruh delivého pletiva, takže druhotne hrubnú. Typické pre borovicorasty sú ich listy, ktoré sú väčšinou (ale nie vždy) ihlicovité alebo šupinovité s hrubou pokožkou a kutikulou. Prieduchy listov sú často vnorené do listu, čím sa znižuje odpar vody. Listy môžu vyrastať po jednom (napr. smrek), po dvoch až piatich (borovica), alebo vo zväzočku na typických skrátených konároch nazývaných brachyblasty (napr. smrekovec). Väčšina druhov je stálezelených, niektoré druhy sú opadavé (napr. smrekovec). Vytvárajú jednopohlavné (monoecické) šištice (strobilis). Samčie šištice pozostávajú z tyčiniek s krátkou nitkou na ktorej sú dve puzdrá produkujúce veľké množstvo peľu. Peľ má často 2 vzdušné vaky a je roznášaný vetrom (borovicorasty sú vetroopelivé). Samičia šištica pozostáva z plodných šupín (megasporofyly), ktoré sa rozdeľujú na podporné šupiny a semenné šupiny. Na nich sa nachádzajú 2 vajíčka, ktoré sú svojím otvorom (mikropilárny otvor) obrátené smerom k stredovej ose šištice. Vajíčko (ovulum) obsahuje samotnú vajcovú bunku (oosféru), 2 podporné bunky (archegóniá) a vyživovacie pletivo (primárny endosperm). Po opelení prerastá peľové zrnko pomocou peľového vrecúška cez otvor nazývaný mikropile do vajíčka. V ňom vznikajú dve neobrvené spermatické bunky, z ktorých jedna oplodní oosféru a druhá zaniká. Po oplodnení sa zo samičej šištice vytvorí väčšinou drevnatá šiška. Šišky sú veľmi variabilné a sú dôležitými znakmi pri rozoznávaní jednotlivých druhov.
Ihličnany (Pinopsida) sú evolučne veľmi staré dreviny, známe už od konca karbónu. Majú úzke ihlicovité alebo šupinovité listy s hlboko vnorenými prieduchmi a hrubou kutikulou. Zvyčajne ide o jednodomé rastliny tvoriace drevnaté šišky. Sú však citlivé na znečistenie prostredia.
Čeľaď borovicovité (Pinaceae): Zástupcovia majú vo svojich pletivách živicové kanáliky. Ihličnany z rodu Pinus (borovice) majú ihlice vo zväzočkoch. Bežné sú borovica lesná (Pinus sylvestris), borovica čierna (Pinus nigra), borovica limbová (Pinus cembra) a borovica horská (kosodrevina, Pinus mugo). Významným druhom je jedľa biela (Abies alba). Z rodu smrekov je pôvodný smrek obyčajný (Picea abies). Jediným opadavým ihličnatým stromom je smrekovec opadavý (Larix decidua). Z nepôvodných druhov sa pestuje duglaska tisolistá (Pseudotsuga menziesii).
Čeľaď cyprusovité (Cupressaceae): Vždyzelené stromy a kry rozšírené po celom svete. Listy sú v dospelosti najčastejšie šupinovité. Z domácich druhov je významná borievka obyčajná (Juniperus communis) s dužinatou šuškou (galbulus). Z nepôvodných je častá tuja západná (Thuja occidentalis).
Čeľaď tisovité (Taxaceae): Zahŕňa dvojdomé dreviny. Kľúčovým zástupcom je tis obyčajný (Taxus baccata) s červeným sladkým mieškom obklopujúcim semeno.
Čeľaď tisovcovité (Taxodiaceae): Zahŕňa reliktné stromy. Absolútnym výškovým rekordérom je sekvoja vždyzelená (Sequoia sempervirens).

Krytosemenné rastliny (Angiospermae)
Krytosemenné rastliny (Angiospermae) patria evolučne najmladšie a najvyššie organizované rastliny (približne 220 000 druhov). Vajíčka a následne aj semená nie sú voľne vystavené, ale sú ukryté v semenníku, z ktorého po dozretí vzniká plod (carpus). Plody, ktoré môžu byť suché alebo dužinaté, chránia semeno a aktívne sa zúčastňujú na jeho rozširovaní.
Krytosemenné rastliny vytvorili pestrofarebné a voňavé kvety s nektárom, vďaka čomu na opeľovanie využívajú živočíchy a hmyz (zoogamia), čo je oveľa efektívnejšie ako vietor. Unikátom tejto skupiny je dvojité oplodnenie - jedna spermatická bunka oplodní vajíčko (vzniká embryo) a druhá splynie s polárnymi jadrami, čím vzniká triploidné vyživovacie pletivo (endosperm). V semenníkoch je buď veľa vajíčok, potom sú plody mechúriky, al. obsahujú iba jedno vajíčko, potom je plodom kôstkovica.
Pre magn. majú na zárodku dva protistojné klíčne listy alorízia /koreňová sústava pozostáva z hlavného koreňa a bočných koreňov/ v stonkách sú cievne zväzky usporiadané do kruhu, listy majú perovitú alebo dlaňovitú žilnatinu, kvety bývajú 5-početné al. výrazne predĺžené kvetné lôžko, široké tyčinky, plodom je najčastejšie mechúrik al. nažka, plodstvo tvarom pripomína šišku.
Pôvodne boli kvetné časti usporiadané v závitnici-acyklické kvety. Postupne vznikali kvety, kde boli niektoré časti v kruhu a niektoré v závitnici. Napokon vznikli cyklické kvety, kt. plodolisty vývojovo starých magnóliorastov mali tvar listu predĺžene zloženého a s okrajmi iba k sebe priloženými. Neskôr zrástli do piestika, tvoreného dutým semenníkom, na vrchole s bliznou. V semenníku sú vajíčka. Po ich oplodnení vznikajú semená a zo semenníkov vznikne plod, kt. uzatvára a chráni semená.
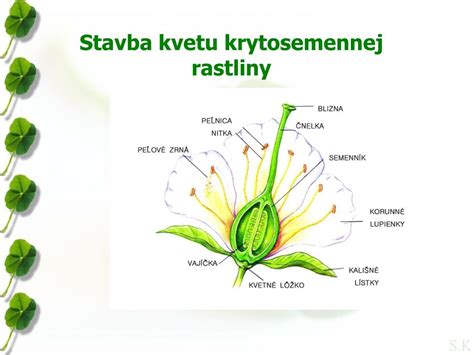
Magnóliorasty (Magnoliophyta)
Moderné fylogenetické štúdie DNA (systémy APG - Angiosperm Phylogeny Group; napr. APG II (2003), APG III (2009) a APG IV (2016)) ukázali, že kým jednoklíčnolistové rastliny tvoria jednu ucelenú vývojovú vetvu (sú monofyletické), obrovská historická skupina dvojklíčnolistových rastlín pochádza z viacerých rôznych vývojových línií (sú polyfyletické). Najnovšie sa preto namiesto podtried používa pojem vetva. Taktiež sa upúšťa od termínu magnóliorasty (Magnoliophyta), ktorý nahrádza pojem krytosemenné rastliny (Angiospermatophyta).
Systém APG rozdeľuje krytosemenné rastliny na niekoľko hlavných monofyletických vetiev:
- Bazálne krytosemenné rastliny - evolučne najprimitívnejšie druhy s jednoduchšími cievicami a peľom len s jedným otvorom (napríklad lekno). Tvoria už len 0,5 % dnešných druhov krytosemenných rastlín.
- Magnoliidová vetva - zachovávajú si niektoré primitívne znaky (peľ s jedným otvorom), no vyznačujú sa tvorbou éterických olejov (napríklad magnólie, vavríny, čierne korenie). Tvoria asi 2,5 % druhov.
- Jednoklíčnolistové rastliny - táto skupina ostala zachovaná ako monofyletická vetva (napríklad trávy, obilniny, orchidey, cibule). Tvorí asi 28 % druhov.
- Pravé dvojklíčnolistové rastliny (eudikotyledony) - ich hlavným moderným identifikačným znakom je trikolpátny peľ (peľové zrno s tromi klíčiacimi otvormi). Táto obrovská skupina sa ďalej delí na menšie vetvy.
Čeľaď Magnóliovité (Magnoliaceae)
Magnólia veľkokvetá je vždyzelený strom dosahujúci výšku 25 m. Má vajcovité červené plody dlhé do 10 cm. Tento druh sa vyskytuje na brehoch riek a vlhkých miestach juhovýchodu USA.
Čeľaď Leknovité (Nymphaeaceae)
Lekno biele.
Čeľaď Iskerníkovité (Ranunculaceae)
Rastliny s jednoduchými kvetmi, ich mliečnice obsahujú alkaloidy. Väčšina druhov je jedovatých, niektoré sa používajú na prípravu liečiv. Rastú prevažne v chladnom podnebí. Iskerník prudký. Pre magn. majú na zárodku dva protistojné klíčne listy alorízia /koreňová sústava pozostáva z hlavného koreňa a bočných koreňov/ v stonkách sú cievne zväzky usporiadané do kruhu, listy majú perovitú alebo dlaňovitú žilnatinu, kvety bývajú 5-početné al. výrazne predĺžené kvetné lôžko, široké tyčinky, plodom je najčastejšie mechúrik al. nažka, plodstvo tvarom pripomína šišku.
Čeľaď Makovité (Papaveraceae)
Bohatá na mliečnice. Mak siaty - plodom je mnohosemenná tobolka (makovica). Narezávaním nezrelých toboliek sa získava surové ópium, ktoré obsahuje mnohé alkaloidy (morfín, kodeín, papaverín).
Čeľaď Kapustovité (Brassicaceae)
Rastliny obsahujú idioblasty, ktoré im dávajú kapustovitú horkastú chuť. Kapusta obyčajná, kapusta hlávková - forma biela alebo červená, kel hlávkový, kel ružičkový, kaleráb, karfiol, repka olejka, biela, polodlhá forma reďkev siata pravá a guľatá reďkev siata čierna, chren dedinský, kapsička pastierska. V pletivách majú idioblasty /bunky obs. štipľavé horčičné silice, kt. spôs. Charakt. chuť kapustovitých rastlín.
Čeľaď Ružovité (Rosaceae)
Pravidelné 5-početné kvety s plochým, vyklenutým, al. prehĺbeným kv. lôžkom a mnohopoč. ako okrasný druh sa pestuje ruža šípová, Nátržník husí, jahoda obyčajná, ostružina černicová a ostružina malinová. Jabloň domáca, Hruška obyčajná, Slivka domáca, Slivka trnková, Čerešňa vtáčia, Marhuľa obyčajná, Broskyňa obyčajná.
Čeľaď Bôbovité
Na koreňoch majú hľuzy so symbiotickými nitrifikačnými baktériami a obsahujú veľa bielkovín. Kališné lístky sú zrastené a korunné lupienky voľné, len dva dolné čiastočne zrastené. Fazuľa záhradná, Hrach siaty, Šošovica jedlá (obs. plody dvojnažky.
Rozmnožovanie krytosemenných rastlín (kvitnúcich rastlín) AKTUALIZOVANÉ
Rastliny prechádzajú počas svojho života určitým životným cyklom. Nazývame ho ontogenéza alebo ontogenetický vývin. Prvým je semeno. Keď sa dostane do pôdy, začne klíčiť, vznikne mladá rastlina schopná rásť a fotosyntetizovať. Neskôr začína kvitnúť. Aby zabezpečila zachovanie svojho druhu, má kvety - práve tie sú dôležité pre rozmnožovanie. Napríklad kukurica je jednoročná rastlina. Jej životný cyklus trvá jedno vegetačné obdobie. Mrkva je dvojročná rastlina, ktorá v prvom roku vyklíči zo semena a dostane sa do vegetatívneho štádia, ale až v lete ďalšieho roku kvitne a má semená.
Na rozmnožovanie slúžia semená. Na to, aby ich rastlina mala, potrebuje špeciálny orgán - kvet. V ňom sa nachádzajú samčie i samičie pohlavné orgány. Samčie sa nazývajú tyčinky a samičí sa volá piestik. Kvet v tyčinkách vyrába peľ. Ten včela, vietor alebo aj voda prenesú z tyčinky na piestik. Potom nasleduje druhý krok - oplodnenie, splynutie samčej a samičej pohlavnej bunky. Ako jediné vznikajú redukčným delením - meiózou. Pri tomto delení sa znižuje počet chromozómov na polovicu. Je to dôležité, pretože pri oplodnení splynie samčia a samičia pohlavná bunka, vznikne jedna dôležitá bunka, ktorú nazývame zygota. Z tejto jednej jedinej bunky vznikne rastlina. V jadre zygoty je uložená genetická informácia, kde je zapísané, ako bude rastlina vyzerať, aké bude mať vlastnosti, kvety, korene, listy. Po vzniku sa zygota začína hneď deliť. V priebehu embryonálneho vývinu sa postupne vyvíja celá klíčna rastlina, ktorá vyklíčila zo semienka. Teda zo zygoty sme získali klíčnu rastlinu. Bunky schopné deliť sa nájdeme v meristéme stonky, kde sa ďalej delia a zvyšuje sa ich počet - prebieha postembryonálny vývin. Tieto jednotlivé bunky sa však od seba odlišujú - špecializujú sa na vykonávanie určitých funkcií. Inak budú vyzerať bunky listu ako bunky koreňa, inak bunky krycích pletív ako bunky vnútri listu. Celá rastlina je tvorená z celých orgánov, pletív a buniek, ktoré sa od seba odlišujú. Všetky bunky rastliny, ktoré sa podieľajú na stavbe rastlinného tela, majú vlastnosť totipotencie. To znamená, že v jadre obsahujú kompletnú genetickú informáciu a za istých podmienok sa táto vlastnosť môže realizovať, čiže z týchto buniek môžeme zase získať kompletné rastliny. I pri tomto procese však existujú výnimky. Jednou z nich sú špeciálne vodivé pletivá, ktoré slúžia na transport látok.
Ľudia už dávno zistili, že rastliny sa môžu rozmnožovať nielen semenami (pohlavne), ale aj vegetatívne (nepohlavne). Pri nepohlavnom rozmnožovaní používame nejakú vegetatívnu časť - či už list alebo odrezky koreňov. Listy plnia dôležitú funkciu, a to že fotosyntetizujú. Prebieha tu premena anorganických látok na organické. Ak list z rastliny odrežeme, snaží sa byť znovu samostatný, znovu získa štatút komplexnej rastliny, a preto dochádza k regenerácii rastliny. Vegetatívne rozmnožovanie listov sa dá robiť aj doma. Treba si odrezať list a ponoriť reznú plochu listovej stopky do prášku, ktorý dostanete v hociktorom kvetinárstve. Obsahuje rastové látky typu auxínov, ktoré napomáhajú tvorbe koreňov - naštartujú proces rizogenézy. List zakorení, ak ho dáte do nádoby s vlhkým pieskom. Po určitom čase môžete listy vybrať a uvidíte, že majú dobre vytvorené korene. Z jedného listu môžeme získať jednu alebo dve rastliny. To sa však ľuďom voľakedy málilo - chceli ich získať čo najviac. Preto sa začalo rozvíjať rozmnožovanie v podmienkach in vitro - rozmnožovanie v malých priestoroch z malých častí. Vezmime si príklad africkej fialky. Keď rozdelíme jej list na osem častí, pri technike in vitro z nej môže vzniknúť až osemdesiat rastlín dokopy.

Potrebujeme materiál, respektíve nejakú časť z neho. Nazývame ho explantát. Je to časť rastliny, ktorú odoberieme. Môže to byť list, časť listu, stonka či jedná jediná bunka alebo protoplast (rastlinná bunka, ktorú chemicky zbavíme bunkovej steny). Podmienky in vitro vytvoríme tak, že sa zvýši regeneračný potenciál rastliny. Základom optimálnych podmienok je príprava živného média. Je ich veľa druhov, najznámejšie je však MS odvodené od mien pánov Murashige a Skooga. Tí už v roku 1962 publikovali článok, v ktorom opísali, ako a z akých zložiek má byť vytvorené živné médium. Pridávame doň aj sacharózu, ktorá je zdrojom energie a uhlíka, ďalej vitamíny, vodu, aminokyseliny, agar (polysacharid získaný z morských rias). Zložkou kultivačného média sú aj rastové látky, ktoré si dokáže vytvoriť i samotná rastlina. Musíme mať k dispozícii aj špeciálny prístroj na sterilizáciu. Očkovanie, rozmnožovanie a nakladanie na živné médiá sa robí v špeciálnych očkovacích boxoch. Keď si všetko vysterilizujeme a naočkujeme na pripravené médiá, potom pestujeme explantáty v špeciálnych kultivačných komorách. Sú to miestnosti, kde sú naukladané police a dôležité je, že v nich musíme nastaviť vhodné fyzikálne podmienky na kultiváciu.
Rastlina je tvorená z veľkého množstva buniek, ktoré sú prispôsobené na vykonávanie nejakej funkcie. Zväčša diferencované bunky budú fotosyntetizovať a aj keď majú jadro, za normálnych podmienok sa nebudú rozmnožovať. Ak však chceme, aby sa rozmnožili, musíme diferencované bunky prinútiť k dediferenciácii. Diferenciácia je proces, keď sa z čohosi jednoduchšieho stáva niečo zložitejšie. Dediferenciácia je opačný proces - bunka sa vo vývine akoby vracia späť, prestáva fotosyntetizovať, dostáva sa do stavu, keď znovu vstupuje do bunkového cyklu a znovu je schopná deliť sa. Pri dediferenciácii môže dôjsť k neorganizovane veľkému kalusu. Kalus je skupina dediferencovaných buniek, ktoré sa opakovane delia a neorganizovane rastú, lebo zabudli spolu komunikovať. Pri dediferenciácii vznikne teda buď kalus, alebo meristematické rastové centrá. Keď chceme regenerovať celú rastlinu formou organogenézy, tak to musíme urobiť v dvoch etapách: v prvej vznikne výhonok a v druhej ho odizolujeme na vhodné kultivačné médiá a regenerujeme koreň. Druhá je cesta somatickej embryogenézy. Je to bipolárna štruktúra, ktorá vznikla zo somatickej bunky, a má dva póly - apikálny meristém koreňa a stonky.
Mäsožravé rastliny sa stali mäsožravé, lebo rastú v pôde, kde sa nachádza málo dusíka a fosforu. Buď tieto rastliny vyhynú alebo sa tomuto nedostatku prispôsobia. Začínajú preto chytať a tráviť korisť. Odlišujú sa rôznymi pascami - najznámejšími sú rosičky okrúhlolisté. Tie rastú aj u nás, ale sú prísne chránené. Na hlavičke majú tekutinu, kde sú voňavé látky a farbivá. Keď sa tam mucha dostane, začne sa metať. List rastliny reaguje tak, že sa začne stáčať a vytvárajú sa tráviace enzýmy, ktoré korisť rozložia. Hrozí, že táto rastlina vyhynie, preto ju rozmnožujeme v podmienkach in vitro. Pri prenose rastlín do pôdy si však musíme dávať pozor, aby sme ich aklimatizovali. Ak by sme to neurobili, veľká časť týchto rastliniek by vyhynula.
tags: #rozmnozovanie #semennych #rastlin #borovicorasty #magnoliorasty