
Minerálna výživa pokusných rastlín: Komplexný prehľad
Výživa rastlín je kľúčová pre ich rast a vývoj, pričom sa striktne líši od výživy živočíchov. Je primárne minerálna (anorganická), čo znamená, že iba minerálne látky plnia úlohu esenciálnych (nenahraditeľných) živín. Organické látky nie sú pre rastliny esenciálne a mnohé z nich sú dokonca toxické.
Rastliny prijímajú z pôdy, vody a vzduchu makroelementy (napríklad dusík, fosfor, draslík) a mikroelementy (železo, zinok). Tieto látky sú nevyhnutné na syntézu sacharidov, bielkovín a lipidov. Pre pestovateľa je nevyhnutné akceptovať základné paradigmy výživy rastlín, ktoré vyplývajú z molekulárnej a energetickej podstaty výživy rastlín.
Základné princípy minerálnej výživy
Minerálnou výživou rastlín sa rozumie príjem minerálnych (anorganických) látok. Zdrojom minerálnej výživy rastlín je pôda (živný roztok) a u vodných rastlín je to voda. Orgánom príjmu je koreň.
Z doteraz známych prvkov sa v rastlinách zistili takmer všetky, ale nie všetky sú pre rastlinu nevyhnutné. Životne dôležité prvky sa nazývajú biogénne prvky. Bez biogénnych prvkov nemôže rastlina normálne žiť a žiadny z týchto prvkov nemôže byť nahradený iným.
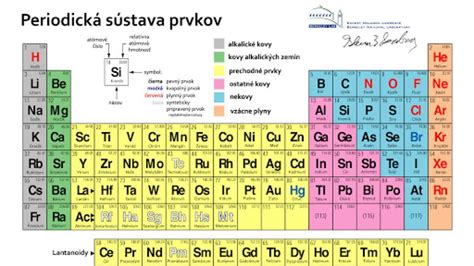
Podľa množstva, v akom sa v rastlinách nachádzajú, delíme prvky na:
- Makroelementy - nachádzajú sa v množstvách od desiatok percenta do stotín percenta. Patria sem C, O, H, N, Si, K, Ca, Mg, Na, Fe, P, S, Al.
- Mikroelementy - nachádzajú sa v množstvách od stotín percenta menej. Patria sem napr. Mn, B, Sr, Cu, Zn, Ba, Ti, Li, Br, Ni, Mo, Co, Cs, Se, Cd, Hg, Ra a iné.
Prvky, ktoré rastlina nevyhnutne potrebuje k životu, nazývame esenciálne, ostatné sú neesenciálne. Zaujímavosťou je, že na rozdiel od živočíchov nie je sodík pre rastliny esenciálny. Esenciálnosť jednotlivých látok sa môže u rôznych druhov mierne líšiť. V prípade niektorých látok potrebných v stopových množstvách (mikroelementy) môže byť ich esenciálnosť sporná.
Fyziologický význam najvýznamnejších prvkov
Uhlík (C)
Uhlík je základom všetkých organických zlúčenín (sušina rastlín obsahuje 30-60 % uhlíka). Do rastlín sa dostáva zo vzdušného CO2 v procese fotosyntézy. Okrem dusíka sa v deficite vyskytuje aj uhlík, ktorého obsah v rastlinnej biomase dosahuje okolo 45 - 50 %. Vo výžive uhlíkom sa predpokladá, že porast je schopný asimilovať až päťnásobne viac CO2, ako to umožňuje jeho reálny obsah v atmosfére.
Kyslík (O)
Kyslík je nevyhnutný prvok, pretože sa zúčastňuje dôležitých procesov dýchania. Pri normálnom (aeróbnom) dýchaní rastliny prijímajú kyslík z atmosféry, pri anaeróbnom dýchaní ho prijímajú zo zlúčenín.
Vodík (H)
Vodík je zložkou vody a organických zlúčenín, je potrebný pri fotosyntéze a rastliny ho prijímajú vodou.
Dusík (N)
Dusík je hlavnou zložkou bielkovín, nukleových kyselín, niektorých rastových regulátorov a vitamínov. Ako zložka bielkovín a tým aj enzýmov sa zúčastňuje na väčšine biochemických reakcií, ktoré umožňujú rastline prežiť. Hoci je dusík najrozšírenejší prvok zemskej atmosféry (78%), je v tejto forme pre rastliny neprijateľný. Korene vyšších rastlín prijímajú dusík v nitrátovej forme (NO3-) alebo ako amónne soli (NH4NO3).
Nezávisle od toho, aké dusíkaté hnojivo bolo do pôdy aplikované (organické, či minerálne - priemyselné), v pôde sa vždy organické N-látky mikrobiálnou aktivitou premenia na minerálne formy dusíka, t. j. amoniak a dusičnan, ktoré sú aj prijímané rastlinami. Akumulácia dusíka v rastlinách je funkciou koncentrácie prijateľného dusíka v pôdnom roztoku, t. j. dusičnanov a amoniaku.
Niektoré rastliny (bôbovité) dokážu pomocou symbiotických baktérií žijúcich na koreňoch prijímať dusík priamo z atmosféry. Dusík sa zabudováva pomocou enzýmov GS (glutamín syntetáza) a GOGAT (glutamát syntáza, nazývaná tiež glutamín-oxoglutarát-aminotransferáza). Výsledkom sú aminokyseliny glutamín (Gln) a kyselina glutámová (Glu).
Deficitné živiny, ku ktorým patrí najmä dusík a uhlík, rozhodujú o produkčnom procese porastu a formovaní úrody. Deficit dusíka je spôsobený najmä tým, že dusík, okrem konštitučnej funkcie (vytvára štruktúru membrán, buniek, pletív a orgánov), zodpovedá za metabolické a syntetické procesy. Tým, že dusík sa využíva na tvorbu vo vode nerozpustných polymérnych látok, je sústavne odčerpávaný na tvorbu štruktúry buniek a zásobných látok (bielkovín). Rovnovážny stav endogénnych zdrojov dusíka je sústavne porušovaný tvorbou polymérnych látok, preto sa vytvára stav deficitu dusíka. V dôsledku uvedeného porast počas celého obdobia vegetácie pozitívne reaguje na prihnojovanie dusíkom.
Fosfor (P)
Fosfor patrí medzi dôležité biogénne prvky, pretože je dôležitou súčasťou nukleových kyselín, fosfolipidov, koenzýmov NAD a NADP a veľmi dôležitý ako zložka ATP. Fosfor prijímajú rastliny koreňmi zo živného média. V pôde sa fosfor nachádza v organickej a anorganickej forme.
Obsah a zásoby fosforu v pôde sú veľmi obmedzené. Vysoká reaktivita aniónov kyseliny trihydrogénfosforečnej spôsobuje ich nízku mobilitu v pôde, a tým aj nedostatočnú prístupnosť pre rastliny. Fosfor v pôde je imobilizovaný chemickou sorpciou vo forme nerozpustných zlúčenín terciárnych fosfátov s Ca, Mg, Fe a Mn.
I napriek uvedenému, v porovnaní s dusíkom, vykazujú rastliny iba mierny deficit fosforu. Osobitosť výživy fosforom je spôsobená predovšetkým tým, že fosfor v rastline vytvára iba malý podiel zásobných polymérnych fosforečných zlúčenín a najmä v jeho unikátnej schopnosti nepretržitej reutilizácie (obnovy) endogénnej zásoby fosforu. V dôsledku reutilizačnej schopnosti sa uskutočňuje kvázi samoregulujúca optimalizácia výživy fosforom. Fosfor vo forme aniónu kys. trihydrogénfosforečnej (H2PO3-) sa aktívne zúčastňuje metabolických a syntetických reakcií, ktoré sú bezprostredne integrované s procesmi fosforylácie a defosforylácie. V dôsledku princípu „sebaobnovy“ vzniká stabilný endogénny rovnovážny stav fyziologickej potreby fosforu. Je mimoriadne dôležité, aby potreby porastu na zdroje fosforu boli saturované v počiatočnej rastovej fáze porastu. Reálne potreby fosforu v porovnaní s dusíkom sú v absolútnych hodnotách desaťnásobne znížené.

Draslík (K)
Draslík (K+) je kofaktorom mnohých enzýmov.
Vápnik (Ca)
Vápnik má význam pri metabolizme rastliny.
Síra (S)
Je mimoriadne opodstatnená aplikácia hnojív s obsahom síry (síran amónny alebo DASA) pri pestovaní olejnín a pšenice pre potravinárske využitie. V prípade pšenice tvorba zásobných lepkových bielkovín (prolamínov a glutelínov) v zrne je bezprostredne podmienená obsahom a frekvenciou výskytu v polypeptidovom reťazci sírnych aminokyselín (cysteínu a metionínu), ktoré poskytujú atóm síry na tvorbu disulfidových väzieb: SH + SH → S - S, ktoré podmieňujú tvorbu sekundárnej, terciárnej a kvartérnej štruktúry bielkovinového agregátu akým je lepok. Podmieňujú nie iba obsah lepku, ale predovšetkým jeho fyzikálno-chemické vlastnosti (pevnosť, rozťažnosť a elasticitu). Kľúčový je enzým cysteín syntáza, ktorý zabuduje síru do aminokyseliny cysteín (Cys).
Príjem živín rastlinami
Orgánom príjmu živín je koreň. Koreňové vlásky majú rozhodujúci význam pre príjem minerálnych živín, lebo ich sorpčná plocha je obrovská. Jednotlivé látky rastlina prijíma selektívne, t. j. prijíma hlavne tie, ktoré pre svoj život najviac potrebuje. Okrem oxidu uhličitého, sú všetky ostatné látky prijímané z pôdy koreňmi, ktoré sú na to dobre prispôsobené.
20 Léčivých A Zázračných Rostlin, Které Byste Měli Mít Doma
Na príjem (ale aj vylučovanie) látok má rastlina pasívne aj aktívne mechanizmy. Medzi pasívne patria kanály, ktoré umožňujú prechod látok v smere od ich vyššej koncentrácie k nižšej. Pri prenikaní iónov do bunky pasívnym transportom sa uplatňujú dve skupiny procesov: intercepcia a kontaktná výmena, objemový tok a difúzia. Objemový tok prebieha vtedy, keď soluty transportuje konvektívny tok vody pôdneho roztoku do koreňa. Veľkosť objemového toku závisí od veľkosti toku vody. Difúzia prebieha vtedy, keď je koncentrácia v pôdnom roztoku vyššia ako v pletivách koreňa. Proces, pri ktorom sa ión pohybuje v smere gradientu elektrochemického potenciálu, pri tomto procese sa spotrebúva voľná energia produkovaná metabolizmom. Dosiahnutá koncentrácia iónov vo vnútornom vakuolárnom roztoku nie je teda výsledkom postupného hromadenia, ale dynamickou rovnováhou medzi tokom do bunky a výtokom do vonkajšieho prostredia. Existencia membrány je podmienkou pre vznik rozdielnych koncentrácií iónov medzi vonkajším a vnútorným prostredím.
Aktívny príjem zabezpečujú prenášače, ktoré využívajú energiu ATP. Využitie ATP môže byť buď priamo prenášačom (primárny transport), alebo nepriame (sekundárny transport). Nepriamy transport prebieha tak, že sa cez tzv. ATP pumpy vylučujú ióny H+ von z buniek a tak vytvárajú elektrický potenciál (napätie) na oboch stranách bunkových membrán. Toto napätie využívajú prenášače na transport látok tak, že dané látky transportujú spoločne s H+.
Cesty pohybu iónov pri jednobunkových rastlinách sú veľmi jednoduché. Transport iónov v xyléme prebieha nielen pôsobením mechanizmov jednoduchého difúzneho toku, ale tiež sériou výmenných procesoch na povrchu stien i povrchovou migráciou na stenách xylémových elementov.

Formy výživy rastlín
Podľa spôsobu, akým rastliny získavajú organické látky, ich rozdeľujeme na:
- Heterotrofné - organické látky si nedokážu samy vyrobiť a prijímajú ich z prostredia.
- Autotrofné - organické látky si vyrábajú z anorganických.
- Mixotrofné - využívajú kombinovanú výživu.
Autotrofia a fotosyntéza
Pojem autotrofia označuje schopnosť organizmu vyrobiť si vlastné organické látky z jednoduchých anorganických zlúčenín (CO₂ a H₂O). Autotrofné organizmy (predovšetkým rastliny) sú tak primárnymi producentmi biosféry, od ktorých priamo či nepriamo závisí zvyšok planéty.
Fotosyntéza je fundamentálny dej, pri ktorom sa svetelná energia Slnka premieňa na stabilnú energiu chemických väzieb organických molekúl. Vedľajším produktom tohto procesu je kyslík. Primárne procesy fotosyntézy vyžadujú prítomnosť svetla, preto bývajú označované ako fotochemická fáza. Základnou úlohou primárnych procesov je premeniť kinetickú energiu fotónov (svetla) na využiteľnú chemickú energiu vo forme molekúl ATP a redukčného činidla NADPH + H⁺. Pre rastliny je dusík prijateľný až po jeho zapravení do pôdy a po uskutočnenej mineralizácii a vzniku anorganických foriem N a P. Na asimiláciu látok v rastlinách sa využíva slnečná energia, ktorá v svetelnej fáze fotosyntézy vytvára dve energeticky bohaté zlúčeniny, ATP a feredoxín. Tieto dve zlúčeniny sa potom využívajú na zabudovanie uhlíka do organických zlúčenín (cukrov) v procese Calvinovho cyklu.
Presne rovnakým spôsobom vykonávajú fotosyntézu aj sinice, ktoré sú prokaryotické organizmy. Zvyšné skupiny fotoautotrofných baktérií (napr. purpurové alebo zelené sírne baktérie) už vykonávajú bezkyslíkatú (anoxygénnu) fotosyntézu. Ako zdroj vodíka na redukciu CO₂ nevyužívajú vodu, ale iné zlúčeniny, najčastejšie sírovodík (H₂S), preto pri ich fotosyntéze nevzniká kyslík, ale uvoľňuje sa síra. Mnohé purpurové baktérie taktiež redukujú NAD⁺ na NADH + H⁺, na rozdiel od rastlín a siníc, ktoré redukujú NADP⁺ na NADPH.
U fotosyntetizujúcich rastlín prebiehajú tieto deje v chloroplastoch na membránach tylakoidov. Hlavným asimilačným farbivom je chlorofyl a, ktorý sa nazýva aj aktívny chlorofyl. Ten prostredníctvom fotooxidácie (uvoľnením elektrónu) premieňa svetlo na chemickú energiu. Ostatné pigmenty (chlorofyl b, karotenoidy) fungujú ako pomocné svetlozberné antény. Okrem chlorofylu sa fotosyntézy zúčastňujú aj iné farbivá ako napríklad karotenoidy a u siníc, červených rias a kryptomonád aj fykobilíny - fykoerytríny a fykocyaníny.
Cyklický a necyklický transport elektrónov
Necyklický transport elektrónov prebieha za účasti oboch fotosystémov. Najprv za účasti PSII dochádza k excitácii chlorofylu v reakčnom centre P680. Zároveň dochádza k aktivácii enzýmu, ktorý rozkladá vodu na protóny, elektróny a kyslík, ktorý uniká do atmosféry. Keďže tento rozpad vody prebieha za prítomnosti svetla, označuje sa termínom fotolýza vody (Hillova reakcia):
\( \ce{2H2O -> O2 + 4H+ + 4e-} \)
Elektróny z fotolýzy vody dopĺňajú chýbajúce miesto v excitovanom P680. Vzbudené elektróny z P680 sa prenášajú ďalej cez cytochróm b6/f na reakčné centrum PSI. Energia, ktorú pri tom elektrón vyžiari, slúži na vznik protónového gradientu, ktorý následne enzým ATP syntáza využíva na syntézu ATP. Tomuto procesu hovoríme necyklická fotofosforylácia.
Cyklický transport elektrónov prebieha len za účasti PSI. Začína pohltením svetelnej energie molekulou chlorofylu v reakčnom centre P700. Z chlorofylu sa tým uvoľnia elektróny, ktoré zachytí oxidačno-redukčný enzým feredoxín. Z neho sa elektróny prenášajú späť na chlorofyl cez plastocyanín a cytochrómový komplex b₆/f. Keďže pri tomto procese vykonávajú elektróny cyklus (chlorofyl - feredoxín - oxidačno-redukčné enzýmy - chlorofyl), označujeme tento dej ako cyklická fotofosforylácia. Cyklický transport elektrónov predstavuje alternatívnu dráhu, ktorú chloroplast volí v prípade nedostatku NADP a ATP.
Tmavá fáza fotosyntézy (Calvinov cyklus)
Sekundárne procesy priamo nevyžadujú prítomnosť svetla, a sú preto označované ako tmavá alebo termochemická fáza fotosyntézy. Počas týchto procesov dochádza k fixácii CO₂ a vzniku sacharidov. Zdrojom energie na túto premenu je ATP a redukovadlom NADPH + H⁺. Pomenovanie tmavá fáza nie je úplne šťastné, pretože to evokuje, že tento dej prebieha v tme. Nie je to však pravda. Hoci je tento dej nezávislý od priameho slnečného žiarenia, vyžadované sú pri ňom východiskové produkty z primárnych procesov fotosyntézy − ATP a NADPH. Cesta fixácie CO₂ týchto rastlín sa nazýva Calvinov cyklus. Reakcie možno rozdeliť do 3 fáz:
- Karboxylačná fáza − primárnym akceptorom CO₂ je ribulóza-1,5-bisfosfát (RuBP), čím vzniká nestabilná 6-uhlíkatá molekula, ihneď sa hydrolyzujúca na 2 3-uhlíkaté (preto C3) molekuly kyseliny 3-fosfoglycerovej; reakciu väzby CO₂ na RuBP katalyzuje enzým rubisco (ribulóza-1,5-bisfosfát karboxyláza/oxygenáza), ktorý v listoch tvorí 30-50 % všetkých prítomných bielkovín; katalytická aktivita rubisco je podmienená prítomnosťou Mg²⁺.
- Redukčná fáza − kyselina 3-fosfoglycerová je redukovaná na glyceraldehyd-3-fosfát za spotreby ATP a NADPH + H⁺.
- Regeneračná fáza − regenerácia akceptora RuBP.
Fotorespirácia
Fotorespirácia je proces, kedy rastlina spotrebováva O₂ a produkuje CO₂. Je to teda opačný proces k fotosyntéze. Od dýchania sa odlišuje tým, že pri fotorespirácii sa energia v podobe ATP a redukovadlo NADPH spotrebováva. Prvotnú reakciu fotorespiračnej kaskády paradoxne zahajuje taktiež enzým rubisco, avšak ako substrát používa O₂ namiesto CO₂ (preto sa nazýva karboxyláza (viaže CO₂)/oxygenáza (viaže O₂)).
V oxygenázovej reakcii reaguje O₂ s RuBP a vzniká kyselina 3-fosfoglycerová a 2-fosfoglykolát, ktorý sa ďalej enzymaticky metabolizuje až na CO₂. Tieto deje prebiehajú čiastočne aj v peroxizómoch a mitochondriách za spotreby ATP a NADPH.
Úloha fotorespirácie nie je uspokojivo vysvetlená, ale predpokladá sa, že by mohla slúžiť k ochrane rastlín pred nežiadúcimi účinkami nadmerného slnečného žiarenia pri súčasnej absencii CO₂. Pri fotorespirácii zároveň vznikajú aj prekurzory pre syntézu niektorých aminokyseliny (glycín, serín). Možná je aj hypotéza, že afinita O₂ k enzýmu rubisco vznikla ešte v časoch, kedy atmosféra obsahovala veľmi málo O₂, a teda neexistovala negatívna selekcia proti tomuto fenoménu.
C4 a CAM rastliny
Anatómia listov C4 rastlín sa odlišuje od C3 rastlín a dochádza aj k priestorovému oddeleniu fixácie CO₂ (bunky mezofylu) a jeho redukcie na glukózu (bunky pošiev cievnych zväzkov). Takto si pre enzým Rubisco udržiavajú umelo obrovskú lokálnu koncentráciu CO₂, čím úplne zabránia fotorespirácii a zachovajú si masívne výnosy cukrov. Primárnym akceptorom CO₂ v mezofyle listov je fosfoenolpyruvát (PEP), pričom jeho karboxyláciou vzniká 4-uhlíkatá (preto C4) kyselina oxaloctová, ktorá sa ďalej pomocou NADPH + H⁺ redukuje na malát. Malát sa transportuje do pošiev cievnych zväzkov, kde sa dekarboxyluje a vzniknutý CO₂ vstupuje do Calvinoho cyklu.
Sukulenty a kaktusy z extrémnych púští šetria vodou tak, že okrem priestorového oddelenia fixácie a redukcie CO₂ oddelili procesy aj časovo. Tieto tzv. CAM rastliny (angl. Crassulaceae Acid Metabolism) predstavujú variant C4 rastlín, ktoré majú cez deň zatvorené prieduchy, aby predišli stratám vody. Zároveň však neprijímajú ani CO₂. CAM rastliny teda prijímajú CO₂ v noci a v cytoplazme ho pomocou PEP-karboxylázy viažu na fosfoenolpyruvát (PEP). Vzniknutá kyselina oxaloctová sa pomocou NADPH + H⁺ redukuje na malát. PEP a NADPH + H⁺ sa tvoria procesom glykolýzy, takže súčasne sa znižuje obsah škrobu v chloroplastoch. Malát sa transportuje do vakuoly a okysľuje bunkovú šťavu.
Heterotrofia
Saprofytické rastliny odoberajú organické látky z odumretých tiel iných organizmov, čím ich mineralizujú na jednoduché látky (CO₂, H₂O, NH₃). Parazitné organizmy (holoparazity) odoberajú živiny inému živému organizmu - hostiteľovi. Do jeho tela vnikajú pomocou premenených koreňov, takzvaných prísaviek (haustórií). Poloparazity (hemiparazity) majú zelené listy a normálne fotosyntetizujú. Svojho hostiteľa využívajú len ako zdroj vody a minerálov.
Mixotrofia
Mixotrofia je špecializovaný spôsob výživy spájajúci autotrofiu a heterotrofiu. Rastliny normálne fotosyntetizujú, no popritom aktívne prijímajú iné organické živiny z tiel živočíchov. Tento jav je typický pre mäsožravé rastliny. Žijú v drsných a podmáčaných biotopoch (rašeliniská), kde je v pôde pre absenciu užitočných baktérií kritický nedostatok dusíka. Deficit dusíka pre tvorbu bielkovín si preto kompenzujú lovom hmyzu pomocou premenených listov.
Jednotlivé druhy si vyvinuli rôzne typy pascí. Bublinatka (Utricularia) loví vo vode pomocou malých komôrok, ktoré hmyz pri podráždení bleskovo nasajú podtlakom. Rosička (Drosera) má na listoch lepivé trichómy, na ktoré sa hmyz prilepí, list sa zvinie a vylúči tráviace enzýmy. Mucholapka (Dionaea) chytá korisť aktívnym, mimoriadne rýchlym zaklapnutím dvoch polovíc listu, zatiaľ čo krčiažnik (Nepenthes) láka hmyz do pasívnych pascí v tvare krčahov naplnených tráviacimi šťavami. Mixotrofia je veľmi bežná aj u jednobunkových organizmov a rias.

Faktory ovplyvňujúce fotosyntézu a výživu
- Svetlo - primárny zdroj energie. Najdôležitejšie je svetlo v červenej a modrofialovej časti spektra (okolo 680 a 700 nm). Rastlina dokáže zo svetla, čo na ňu dopadne, využiť asi 2 %. Ostatné svetlo sa odráža, alebo prepúšťa. Pri priveľmi silnom žiarení sa fotosyntéza zastavuje kvôli poškodeniu buniek reaktívnym kyslíkom.
- Oxid uhličitý (CO₂) - kľúčový zdroj uhlíka. Z jedného gramu CO₂ sa vytvorí asi 0,5 g sušiny. Bežné rastliny dokážu zvyšovať efektivitu fotosyntézy priemerne až do koncentrácie 0,05 % CO₂ vo vzduchu.
- Voda (H₂O) - priamy zdroj elektrónov a protónov pri fotolýze. Jej nedostatok spôsobí zatvorenie prieduchov a zablokuje prísun CO₂, čím sa fotosyntéza spomalí. CAM rastliny majú niekoľkonásobne nižší prírastok biomasy práve z tohto dôvodu.
- Teplota prostredia - reguluje aktivitu enzýmov. U väčšiny našich rastlín prebieha fotosyntéza v rozmedzí 0−40 °C, no optimum je zvyčajne 25-30 °C.
Význam pôdy a hnojív pre minerálnu výživu
Prirodzeným prostredím, z ktorého rastliny čerpajú živiny, je pôda. Hnojením vytvárame súlad medzi prijateľnými zdrojmi živín v pôde a požiadavkami porastu. Trend pestovateľských technológií zameraný na ekologizáciu agronomických ekosystémov vyžaduje realizovať aplikáciu priemyselných a organických hnojív iba na základe výsledkov diagnostiky potreby porastu na úroveň programovanej úrody a zhodnotenia reálnych zásob prijateľných živín v pôde. Hnojením zabezpečujeme potreby porastu na prijateľné minerálne živiny v požadovanom množstve a chemickej forme.
Organické hnojivá, vrátane maštaľného hnoja a kompostov, je nevyhnutné aplikovať iba na dobre vyhnitých a humifikovaných substrátoch, zbavených ťažkých kovov, pesticídov, mykotoxínov, patogénnych mikroorganizmov a xenobiotík. Uhlíková matrica organických biologických látok prítomných v maštaľnom hnoji, komposte, rastlinných pozberových zvyškoch a zelenom hnojení je nenahraditeľná pre formovanie pôdnej štruktúry, sorpčného komplexu a humusu, ktorý vytvára potenciálne zdroje živín a zodpovedá za mikrobiologickú aktivitu a pôdnu úrodnosť. Každé organické hnojivo, vrátane rastlinných zvyškov po ich mineralizácii, zanecháva významnú uhlíkovú stopu, preto nie je reálne substituovať potreby dusíka iba organickými substrátmi. Jednostranná preferencia organických foriem živín je vo svojej podstate v rozpore s fyziologickou paradigmou minerálnej výživy rastlín.
Pôdna reakcia má priamy vplyv na životaschopnosť rastlín. Pri pH menšom ako 3 a väčšom ako 9 je vážne poškodzovaná protoplazma v koreňových bunkách. Rôzna rýchlosť, ktorou rastlina spotrebováva ióny z určitej soli, určuje fyziologickú reakciu danej soli. Biologický vplyv rastlín na pôdu je podmienený koreňovými výlučkami, ktoré slúžia ako výživa rizosférnych mikroorganizmov. Ich činnosťou dochádza k mobilizácii živín pre rastliny, fixácii vzdušného dusíka, rozkladu ťažko rozpustných zlúčenín a iné. Upevňuje rastlinu v pôde, zásobuje ju minerálnymi živinami a vodou a syntetizuje niektoré pre rastliny veľmi dôležité látky.
Priemyselné hnojivá
Pri hodnotení priemyselných anorganických (okrem močoviny) a organických hnojív i najrôznejších preparátov je žiaduce prísne diferencovať syntetické organické látky a kvázi syntetické minerálne zlúčeniny reprezentované priemyselnými hnojivami, ktoré v žiadnom prípade nepatria do skupiny cudzorodých látok. Priemyselné hnojivá sú vo svojej podstate vo vode dobre rozpustné soli, biodegradovateľné a obsahujúce iba esenciálne biogénne prvky (živiny) pre rastliny.
Ich príprava je realizovaná výlučne technologickými postupmi extrakcie (vylúhovania) živín z prírodných substrátov. Napríklad fosfor sa získava vylúhovaním z deficitných hornín apatitov i fosforitov a čiastočne z kostí živočíchov. Zlúčeniny dusíka sa v minulosti pripravovali výlučne z guana (vtáčí trus) a od 40. rokov minulého storočia z prírodných zdrojov molekulárneho dusíka (N2) z atmosféry, podľa technológie vypracovanej Haberom. Kvázi „výroba“ draselných hnojív spočíva iba v mechanickom spracovaní prírodných sedimentov. Takže príprava K-hnojív nepatrí medzi chemické výrobné technológie, ale ide iba o ťažbu draselnej soli zo sedimentov a o ich mechanickú úpravu, eventuálne vylúhovanie draselnej soli s následnou rekryštalizáciou pri 45 °C.
Priemyselná výroba hnojív sa datuje od roku 1843, kedy Loos (v Anglicku) pripravil prvé priemyselné hnojivo superfosfát, a to z apatitu. Ako rozpúšťadlo použil kyselinu sírovú. V terajšej dobe sa dominantne využíva kyselina trihydrogénfosforečná. V súvislosti s technológiou výroby superfosfátu pokladám za potrebné pripomenúť originálny návrh prof. Ing. Jana Hampla, DrSc. (pracovníka VŠP v Nitre) na substitúciu kyseliny sírovej koloidnou kyselinou kremičitou. Do dnešných dní uplynulo 178 rokov výroby a aplikácie priemyselných hnojív. Za toto obdobie, za predpokladu, že hnojivá boli použité podľa vedeckých zásad, nikde a nikým nebol zaznamenaný žiaden negatívny vplyv narušenia prírodného prostredia či zdravia ľudí a zvierat.
tags: #mineralna #vyziva #pestovanie #pokusnych #rastlim